400-029-0925
400-029-09251IDH基因
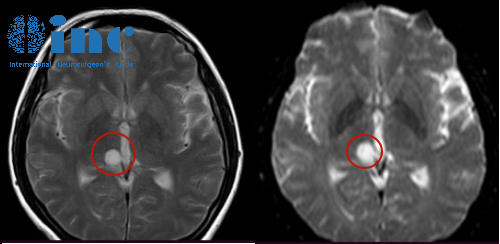
异柠檬酸脱氢酶(isocitrate dehydrogenase,IDH)突变可产生2-羟戊二酸,竞争性控制α-酮戊二酸依赖性脯氨酸羟化酶,升高细胞内缺氧诱导因子-1α,诱导DNA甲基化,于2008年由Parsons等一次利用高通量测序技术发现。为了推广IDH基因检测,Valentina等研制出解吸电喷雾电离质谱分析法,通过检测2-羟戊二酸,3分钟即可判别IDH突变情况,敏感度和特异度均高于80%。与此同时,针对IDH基因的靶向治疗不断涌现,Kim研发出3-氟甲基乙醇,可通过靶向控制IDH1R132H,起到杀伤肿瘤细胞的效果。刚刚FDA批准上市的恩西地平,可逆性控制IDH2突变产生2-羟戊二酸,在急性髓细胞性白血病中已取得成功。诺华公司的IDH1突变控制剂IDH305,也进入神经胶质瘤的II期临床试验。
Schumacher研发了一款针对IDH1突变蛋白的疫苗,IDH1突变蛋白会被表达组织相容复合物II的细胞递呈给CD4细胞,对存在突变的细胞产生特异杀伤。这款疫苗针对III-IV级神经胶质瘤的I期临床试验已于2015年6月完成招募,正在进行。另一个来自杜克大学的治疗II级胶质瘤的PEPIDH1M疫苗也已进入I期临床试验。
2MGMT基因
O6-甲级鸟嘌呤-DNA甲基转移酶(O6-methylguanine-DNA-methyltransferase,MGMT)是一种修复O6-甲级鸟嘌呤烷的酶,而替莫唑胺等烷化剂的作用机制是使O6-甲级鸟嘌呤烷基化,形成DNA交联,从而杀死肿瘤细胞,因而肿瘤表达MGMT是对烷化剂耐药的主要原因。MGMT控制剂O6苄基鸟嘌呤作为卡氮芥增强剂已进入III期临床试验,原理是作为假底物与MGMT结合,起到灭活底物的作用,使肿瘤无法修复O6-甲级鸟嘌呤。但结果显示O6苄基鸟嘌呤对于新诊断胶质母细胞瘤并未好转生存期,且增加了神经毒性。而且Chen等认为肿瘤细胞可由于谷胱甘肽-S-转移酶活性增高,而对O6苄基鸟嘌呤和替莫唑胺耐药。
虽然烷化剂的辅助药物研发遇阻,但新型烷化剂替莫唑胺脂相比替莫唑胺增加了药物脂溶性,使药物进入瘤体内部的浓度增加,对胶质瘤LN229细胞的诱导凋亡作用强于替莫唑胺,且能克服替莫唑胺耐药。另外萝卜硫素能够下调MGMT蛋白表达,联合替莫唑胺可增加其敏感性。但以上两种药物目前停留在体外试验阶段,有待进一步开展临床试验。
3EGFR基因
表皮生长因子受体(epidermal growth factor receptor,EGFR)基因编码一种酪氨酸激酶受体,与配体结合后激活大鼠肉瘤蛋白/磷脂酰肌醇3通路,最终激活底物雷帕霉素靶蛋白,从而促进细胞增殖和能量合成。较常见的突变为截段突变,即外显子2-7框内缺失形成的EGFRVIII重排,不再绑定配体的短胞外区。主要见于原发性胶质母细胞瘤,扩增和过表达随恶性程度增高而增高。
针对EGFR通路已有尼妥珠单抗及其他靶向性替尼单抗,尼妥珠单抗于2004年由美国食品药品监督管理局和欧洲药品管理局双重认证,批准晚期胶质瘤的孤儿用药资格,III期临床试验证实可合适延长无病进展期和总生存期。但一项随机双盲对照II期临床试验显示,另一种抗EGFR单抗西妥昔单抗在复发高级别胶质瘤人群中较少获益,不值得推广。酪氨酸激酶受体控制剂进行了多次III期临床试验,并不成功,而且发现表达KLHDC8A基因可使胶质瘤对吉非替尼和埃罗替尼产生耐药。
看到单抗和酪氨酸激酶受体控制剂的局限性,塞德斯公司研发出针对EGFRVIII重排的疫苗Rindopepimut,III期临床阶段(NCT01480479)已完成,67%接受3个月以上疫苗注射的患者EGFRVIII重排被清除,可合适延长总生存期6个月,已进入IV期临床阶段。另外,Shir在Nature发文显示,体内注入反义RNA修复VIII重排,可使胶质瘤缩小超过40倍。其他RNA如小干扰RNA通过介导转录后基因沉默,可敲除90%目标mRNA,中位生存期延长90%。MiRNA可直接控制EGFR受体转录,下调蛋白激酶B信号表达,在肝癌中已进入I期临床试验(NCT01829971),但目前的载体无法通过血脑屏障,需要寻找其他合适的载体。
4PTEN基因
磷酸脂酶与张力蛋白同源物(Phosphatas and tension homolog,PTEN)通过控制磷脂酰肌醇2磷酸磷酸化为磷脂酰肌醇3磷酸,从而实现对磷脂酰肌醇3/蛋白激酶B信号通路的负性调节,促进细胞凋亡。基因突变、表观遗传沉默或非编码RNA调节可致PTEN纯合子缺失,从而导致激酶S6K1激活,诱发胶质瘤。Liu发现LY-2779964靶向控制S6K1,对PTEN纯合子缺失的肿瘤细胞产生细胞毒作用。Lee发现罗格列酮选择性激活过氧化物酶增殖体活化受体γ,使其结合到PTEN启动子的结合位点,上调PTEN表达,并增强酪氨酸激酶控制剂吉非替尼的抗肿瘤作用。
5P53基因
P53基因编码一种转录因子,诱导缺陷细胞凋亡。P53蛋白主要受鼠双微体蛋白(murine double minute-2,MDM2)的调控,MDM2蛋白通过与P53反式激活结构域结合或催化P53蛋白泛素化降解,使其失活,缺陷细胞就可继续繁殖。Chene等根据P53-MDM2复合体结构,设计了八肽类复合物,无须进一步修饰,即可穿入细胞,激活肿瘤细胞中野生型P53蛋白活性。另外还有硫氧还蛋白、P53蛋白的肽类似物等均通过阻碍P53和MDM2的相互作用治疗表达MDM2和野生型P53的胶质瘤,但这类多肽及类似物容易被代谢,利用率较低。
另外一类非肽类小分子控制剂MI-77301,结构类似于P53,可与MDMD2牢牢结合,竞争性控制P53与MDM2的作用,控制肿瘤细胞生长,生物利用度较高,相比多肽类药物更有研究价值,已进入I期临床试验阶段。
6BRAF基因
鼠类肉瘤病毒同源基因B1(v-rafmurine sarcoma viral oncogene homologB,BRAF),编码一种丝/苏氨酸特异性激酶,是丝裂原活化蛋白激酶通路的重要转录因子,参与细胞生长分化与凋亡。BRAF基因V600E错义突变,在各级别胶质瘤中均可检测到。2011年BRAFV600E控制剂威罗菲尼的上市为胶质瘤带来了的治疗方式。Del等通过威罗菲尼单药使一例儿童颈髓神经节细胞胶质瘤体积明显缩小。Chamberlain回顾分析了4例复发的多形性黄色星形细胞瘤通过威罗菲尼治疗后总生存期延长至8个月,但出现关节炎、皮疹及畏光等不良作用。I/II期临床试验(NCT01006980C)结果显示,另一种BRAFV600E控制剂达拉菲尼对纳入的32例患者中,23例均有应答,且肿瘤明显缩小,这项令人鼓舞的结果也推动了该药联合MEK控制剂的I/II期临床试验,目前试验正在进行。
7TERT启动子突变
端粒酶逆转录酶(telomerase reverse transcriptase,TERT)启动子突变,可使肿瘤细胞获得无限增殖能力。端粒酶作为药物靶点有两个优势,一是很少在正常细胞表达,二是肿瘤细胞中端粒的长度远低于正常细胞,因而选择性控制端粒酶活性可使肿瘤细胞比正常细胞更快死亡。核苷类逆转录酶控制剂(ddG和AZT等)通过和三磷酸单核苷酸竞争与端粒酶的结合位点发挥作用,在治疗T细胞白血病、甲状旁腺癌、食管癌等多种肿瘤中取得成功。
BIBR1532被认为是一种很有前景的端粒酶控制剂,由德国勃林格公司研制,可分别与脱氧核糖核苷酸及DNA引物的位点相结合,并控制端粒酶活性。GRN163L于2006年底由杰龙公司申请开始临床试验,一、二期临床选择在慢性粒细胞性白血病患者中进行,对实体肿瘤的临床试验最近刚刚开始。这类药物的缺点是15%恶性肿瘤细胞可通过端粒延长替代机制维持端粒长度,端粒酶控制剂对这些肿瘤无效。
8ATRX基因
α-地中海贫血/智力低下综合症X染色体连锁基因(alpha thalassemia/mental retardation syndrome X linked,ATRX)编码解旋酶蛋白,通过端粒延长替代机制维持细胞端粒长度。发生于星形细胞瘤、继发性胶母,不发生在少突胶质细胞瘤,多伴有IDH和TP53突变,与1p/19q共缺失和TERT启动子突变排斥。AZ20是一次报道体内合适的选择性ATR激酶控制剂,可修复ATRX突变基因,恢复端粒功能。目前针对ATRX基因的靶向药物较少,尚缺乏相关的临床试验。
- 文章标题:胶质瘤的潜在治疗靶点
- 更新时间:2021-11-05 15:38:27