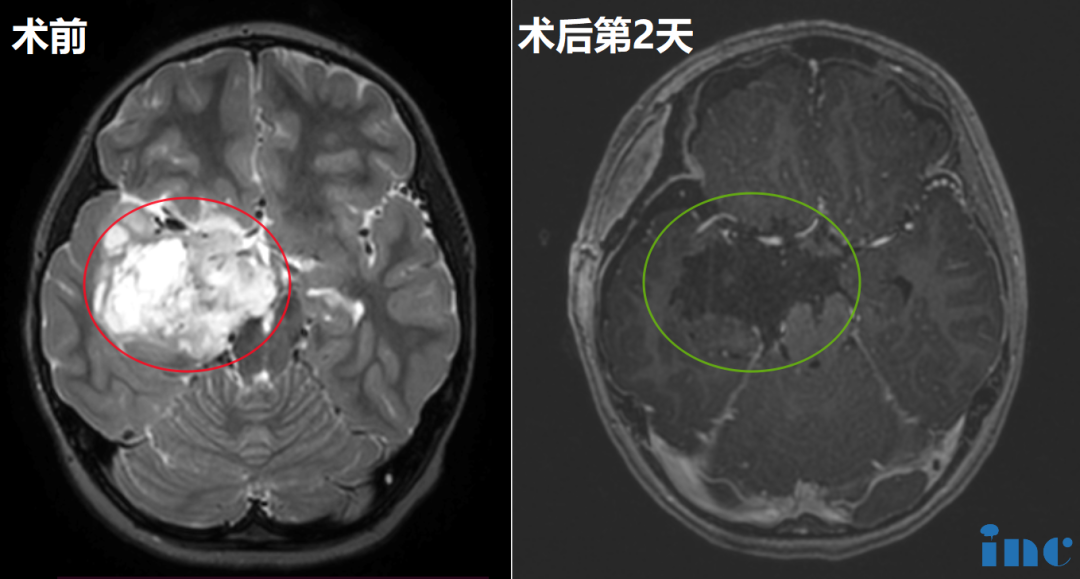
400-029-0925
400-029-0925INC国际儿童脑瘤大咖、世界小儿神经系统知名杂志《Child´s Nervous System》现任主编Concezio Di Rocco教授发表研究《Tis21Knock-Out Enhances the Frequency of Medulloblastoma in Patched1 Heterozygous Mice by Inhibiting the Cxcl3-Dependent Migration of Cerebellar Neurons》(Tis21敲除通过抑制Cxcl3依赖性小脑神经元迁移,提高Patched1杂合子小鼠髓母细胞瘤发生率),以下是研究简述。
摘要
小脑颗粒神经元前体细胞(GCPs)位于小脑外颗粒层(EGL),其增殖失控可导致髓母细胞瘤。为研究GCPs的肿瘤转化过程,我们将低发髓母细胞瘤的Patched1杂合子(Patched1+/-)小鼠与Tis21基因缺失小鼠杂交,建立新的髓母细胞瘤模型。Tis21过表达可抑制GCPs增殖并诱导其分化,而在人髓母细胞瘤中其表达下调。双敲除小鼠髓母细胞瘤发生率显著升高,且出现由前肿瘤性GCPs形成的EGL增生性病变。Tis21缺失不影响GCPs增殖,但抑制其分化,并显著削弱其离开EGL的内在迁移能力。该迁移缺陷可能是肿瘤发生的关键步骤:滞留于增殖微环境内的GCPs更易转化。通过全基因组分析,我们发现趋化因子Cxcl3是Tis21的直接靶基因。Tis21缺失小鼠EGL及病变中Cxcl3表达下调;向小脑切片外源补充Cxcl3可挽救Tis21-/-GCPs的迁移缺陷,并显著减小增生病变面积。鉴于Tis21可激活Cxcl3转录,本研究结果提示Tis21通过Cxcl3诱导GCPs迁移,Cxcl3或成为髓母细胞瘤治疗的新靶点。
材料与方法
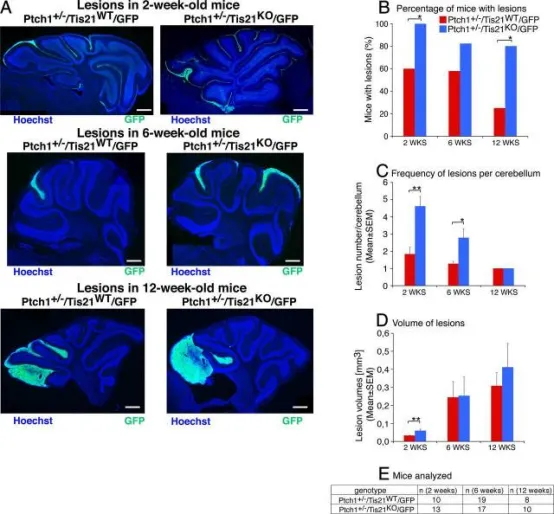
在Patched1杂合背景下敲除Tis21可显著提高出现增生病变的小鼠比例及每侧小脑病变数量。
A:2、6、12周龄Patched1+/-背景下Tis21野生型(Ptch1+/-Tis21WT/GFP)与Tis21缺失(Ptch1+/-Tis21KO/GFP)小鼠小脑矢状切面代表性图像。切片以Hoechst 33258染色,通过Math1增强子驱动的GFP标记增殖性(正常及肿瘤性)GCPs识别病变。标尺470μm。
B–D:2、6、12周龄Ptch1+/-Tis21WT/GFP与Ptch1+/-Tis21KO/GFP小鼠中,出现病变小鼠百分比(B)、每侧小脑病变频率(C)及病变体积(D)。
E:B–D所用小鼠数量。*p<0.05,**p<0.01;B用χ²检验,C、D用Student's t检验。误差线表示SEM。
研究结果
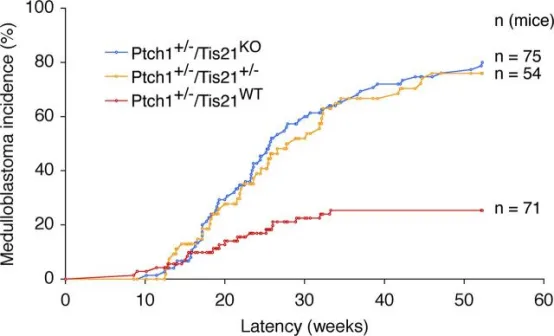
在同时缺失Tis21的Patched1杂合子小鼠中,髓母细胞瘤发生率显著升高。通过将Patched1杂合子与Tis21敲除小鼠至少回交六代,获得Ptch1⁺/⁻/Tis21ᵂᵀ、Ptch1⁺/⁻/Tis21⁺/⁻及Ptch1⁺/⁻/Tis21ᴷᴼ小鼠,随后监测髓母细胞瘤发病情况;与Ptch1⁺/⁻/Tis21ᵂᵀ相比,差异具有统计学意义(p<0.0001,χ²检验)。
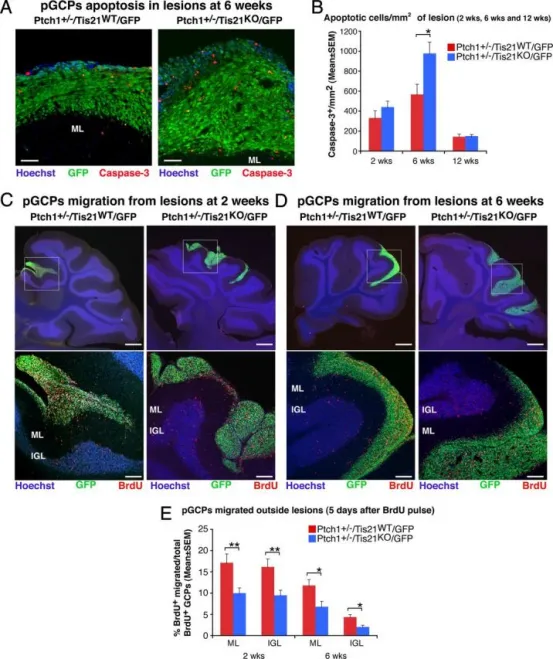
Tis21缺失在Patched1杂合子小鼠中增加前肿瘤性GCP的凋亡并阻碍其迁出病变区
A在6周龄Patched1杂合子小鼠中,利用共聚焦显微镜观察前肿瘤性GCP凋亡(以cleaved Caspase-3阳性细胞标记)。Tis21野生型(Ptch1⁺/⁻/Tis21ᵂᵀ/GFP)与Tis21缺失(Ptch1⁺/⁻/Tis21ᴷᴼ/GFP)小鼠的弥散性增生病变中,前肿瘤性GCP为GFP⁺;分子层(ML)及其边界由Hoechst 33258染色显示。标尺50μm。
B以每个病变区GFP⁺区域为界限,统计整个病变区内cleaved Caspase-3⁺细胞数,计算为每mm²的平均值±SEM;每只小鼠至少取5张非相邻矢状切片。所用小鼠数量见图2E。
C、D在2周或6周龄Patched1杂合子小鼠中,利用BrdU标记迁出病变区的GCP(红色)。Tis21野生型(Ptch1⁺/⁻/Tis21ᵂᵀ/GFP)与Tis21缺失(Ptch1⁺/⁻/Tis21ᴷᴼ/GFP)小鼠的矢状切片用Hoechst 33258复染,显示ML或内颗粒层(IGL)。上方大图框内病变区(标尺410μm)在下方面板以高倍放大(标尺100μm)。
E以BrdU脉冲后5天计算迁出病变区的GCP比例:将邻近病变区的ML或IGL中BrdU⁺细胞数,除以病变区、ML和IGL内BrdU⁺细胞总数,并以百分比±SEM表示。2周龄小鼠每基因型3只,6周龄小鼠每基因型至少4只;每只小鼠所有病变均纳入分析。B与E图中*p<0.05,**p<0.01,Student's t检验。
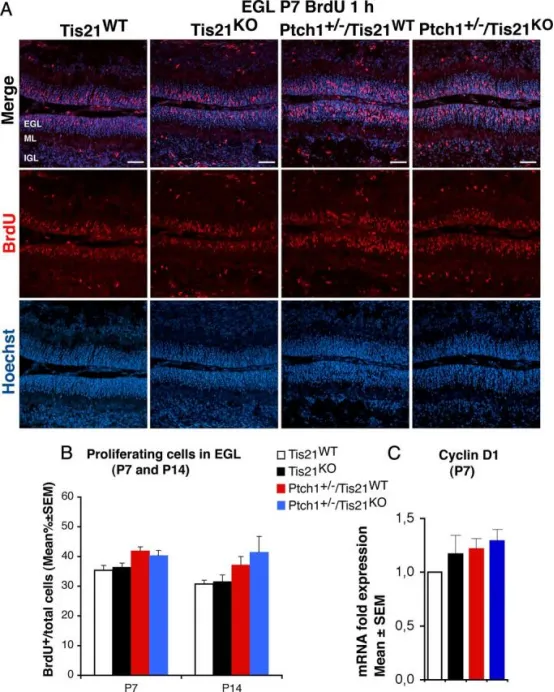
Tis21缺失对Patched1杂合子或野生型小鼠外颗粒层GCPs增殖无影响
AP7龄四种基因型小鼠外颗粒层(EGL)内GCPs进入S期的代表性共聚焦图像,以1 h短脉冲BrdU标记BrdU⁺细胞;切片用Hoechst 33258复染,标示EGL及其所含颗粒细胞与前体细胞。标尺50μm。
BP7及P14龄各基因型小鼠EGL内进入S期的GCPs定量:以BrdU⁺细胞数与Hoechst 33258标记总细胞数之比(BrdU标记指数)表示,数据为平均值±SEM。每基因型3只小鼠。
C实时荧光定量PCR检测P7小鼠分离GCPs的mRNA表达变化,以野生型设为1。数据为3次独立实验的平均值±SEM,采用TBP和GAPDH进行归一化。
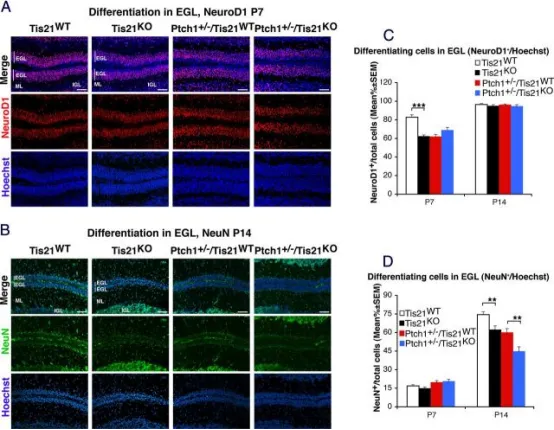
在Patched1杂合子或野生型小鼠中敲除Tis21可抑制外颗粒层(EGL)内颗粒神经元前体细胞(GCPs)的分化。
A、B代表性共聚焦图像显示四种基因型小鼠EGL内GCPs的分化标志物表达:P7小鼠NeuroD1(A)及P14小鼠NeuN(B)。切片以Hoechst 33258复染,用于显示EGL、分子层(ML)和内颗粒层(IGL)。标尺50μm。
C、D定量分析:在P7和P14小鼠EGL内,分别计算NeuroD1⁺(C)或NeuN⁺(D)细胞数与Hoechst 33258标记总细胞数之比,以平均值±SEM表示(每基因型n=3)。**p<0.01,***p<0.001,Student’s t检验。
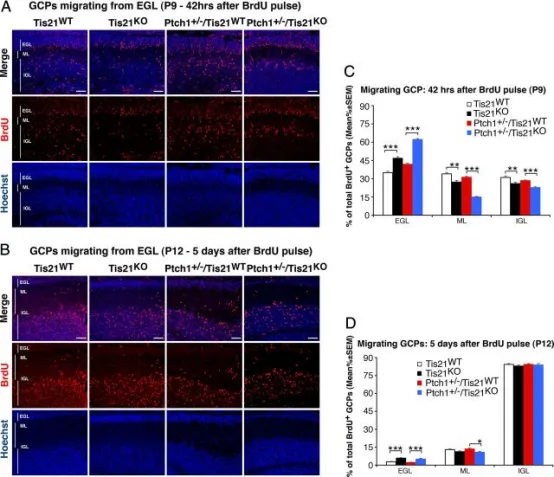
在Patched1杂合或野生型背景下敲除Tis21会损害GCPs从外颗粒层(EGL)向分子层(ML)和内颗粒层(IGL)的迁移。
A、B为典型共聚焦图像,显示在P7小鼠注射BrdU后42小时(A)或5天(B)内,Tis21缺失或野生型小鼠(Patched1杂合或野生型背景)中迁出EGL的BrdU⁺GCPs。切片用Hoechst 33258复染,以显示ML和IGL。标尺50μm。
C、D为定量结果:C为P7小鼠注射BrdU后42小时,D为注射后5天,计算EGL、ML或IGL中BrdU⁺细胞占总BrdU⁺细胞的百分比(平均值±SEM)。每基因型3只小鼠。*p<0.05,**p<0.01,***p<0.001,Student’s t检验。
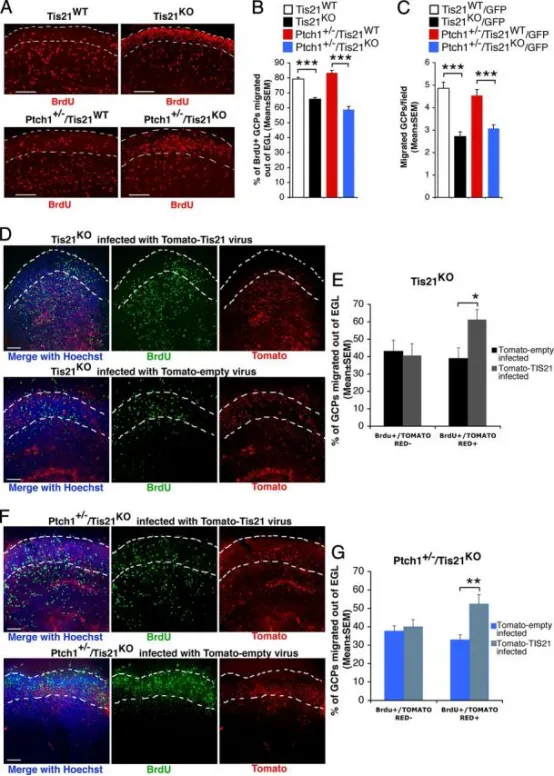
Tis21缺失小鼠GCP迁出外颗粒层(EGL)障碍可被Tis21在脑片培养中挽救
AP7小鼠小脑器官型切片代表性共聚焦图,显示BrdU⁺GCP位于EGL内或已迁出EGL(白虚线标界)。切片经6 h BrdU脉冲标记后,继续培养72 h,固定并以抗BrdU抗体染色。标尺100μm。
BBrdU⁺细胞中位于EGL外者所占百分比(平均值±SEM,3次实验,每次3只小鼠)。***p<0.001,Student’s t检验。
C改良Boyden小室检测纯化GCP的内在迁移能力。用FACS从P7小脑分选GFP⁺GCP,置于上室,计数迁移至膜下侧的GFP⁺细胞。数据为3次独立实验的平均值±SEM,每次计数20视野/孔(≥2孔/实验)。Tis21缺失(Patched1野生型或杂合型)GCP迁移数显著低于Tis21野生型。***p<0.01,Student’s t检验。
D–GTis21逆转录病毒在脑片培养中挽救Tis21⁻/⁻GCP迁移缺陷
D、FP7 Tis21⁻/⁻(D)或Ptch1⁺/⁻Tis21⁻/⁻(F)小鼠小脑切片感染Tomato-Tis21或Tomato-empty逆转录病毒,6 h BrdU脉冲标记,继续培养72 h,固定并以抗BrdU抗体染色。标尺100μm。
E、G在Tis21⁻/⁻(E)或Ptch1⁺/⁻Tis21⁻/⁻(G)切片中,计算BrdU⁺/Tomato⁺或BrdU⁺/Tomato⁻细胞位于EGL外的百分比,并以总BrdU⁺细胞为分母。BrdU⁺/Tomato⁻细胞无显著变化,证实感染特异性。数据为3次实验平均值±SEM,每次3只小鼠。*p<0.05,**p<0.01,Student’s t检验。
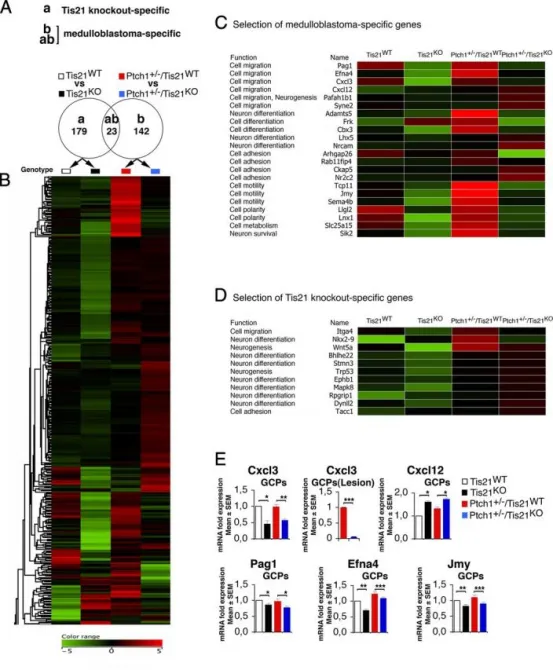
Patched1杂合/Tis21缺失模型中的髓母细胞瘤特异性基因表达
A维恩图所示:对P7小鼠GCPs进行芯片分析,仅在与野生型(Tis21ᴷᴼvs Tis21ᵂᵀ)的比较(a)中鉴定出179个Tis21敲除特异性差异基因;同样,在与Patched1杂合/Tis21野生型(Ptch1⁺/⁻/Tis21ᴷᴼvs Ptch1⁺/⁻/Tis21ᵂᵀ)的比较(b)中鉴定出165个髓母细胞瘤特异性差异基因;其中23个基因在两项比较中均显著差异。
B采用层次聚类算法(欧氏距离,质心法)对两项比较中共344个差异表达基因进行排序;颜色表示相对中位数的高(红)到低(绿)表达变化。
C22个髓母细胞瘤特异性基因参与细胞迁移、分化或黏附过程。
D11个Tis21敲除特异性基因参与细胞迁移或分化过程。
E实时PCR验证5个神经元髓母细胞瘤特异性基因在P7 EGL的GCPs或6周龄病变(GFP⁺)中的差异表达,以野生型小鼠设为1;数据为3次独立实验的平均值±SEM,以TBP和GAPDH归一化。*p<0.05,**p<0.01,***p<0.001,Student’s t检验。
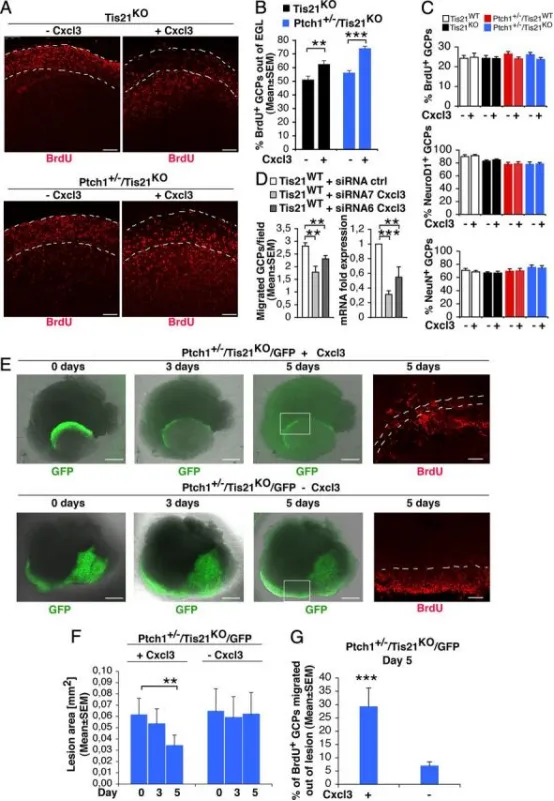
Cxcl3可挽救Patched1杂合/Tis21敲除小鼠GCP迁出EGL或病变区的缺陷,并缩小病变面积
ATis21缺失/Patched1野生型或杂合型小鼠小脑器官型切片经±Cxcl3处理后,BrdU⁺GCP位于EGL内或外(白虚线)的代表性共聚焦图。标尺100μm。
B迁出EGL的BrdU⁺GCP定量:P7切片经±Cxcl3培养5 d(t0加BrdU 18 h),固定后BrdU免疫染色。数据为3次实验(每基因型3只小鼠)BrdU⁺细胞外/总比值的平均值±SEM。**p<0.01,***p<0.001,Student’s t检验。
CCxcl3处理后EGL内增殖(BrdU⁺)或分化(NeuroD1⁺/NeuN⁺)GCP定量:P7切片±Cxcl3培养48 h(分析前2 h加BrdU),固定后BrdU、NeuroD1、NeuN免疫染色。数据为3次实验(每基因型3只小鼠)各标记细胞占总数百分比的平均值±SEM。
DCxcl3 RNAi抑制GCP迁移(左图)。野生型P7大鼠纯化GCP电转siRNA7-Cxcl3、siRNA6-Cxcl3或对照siRNA,恢复3 h后置于改良Boyden小室上室,16 h后计数迁移细胞。数据为3次独立实验(每实验20视野/孔,≥2孔)的平均细胞数/视野±SEM。右图为相同细胞Cxcl3 mRNA水平(TBP归一化)。**p<0.01,***p<0.001,Student’s t检验。
E4周龄Patched1杂合/Tis21缺失(Ptch1⁺/⁻/Tis21ᴷᴼ/GFP)小鼠脑片±Cxcl3处理后,弥漫性增生病变GFP⁺GCP(绿)及BrdU⁺GCP(红)迁出病变区(白虚线)的代表性图。标尺:GFP 500μm,BrdU 100μm。
F病变面积(F)及迁出病变区的BrdU⁺GCP百分比(G)定量:P42切片±Cxcl3处理5 d(t0加BrdU 18 h),固定后BrdU免疫染色。数据为3次实验(每基因型3只小鼠)的平均值±SEM。**p<0.01,***p<0.001,Student’s t检验。
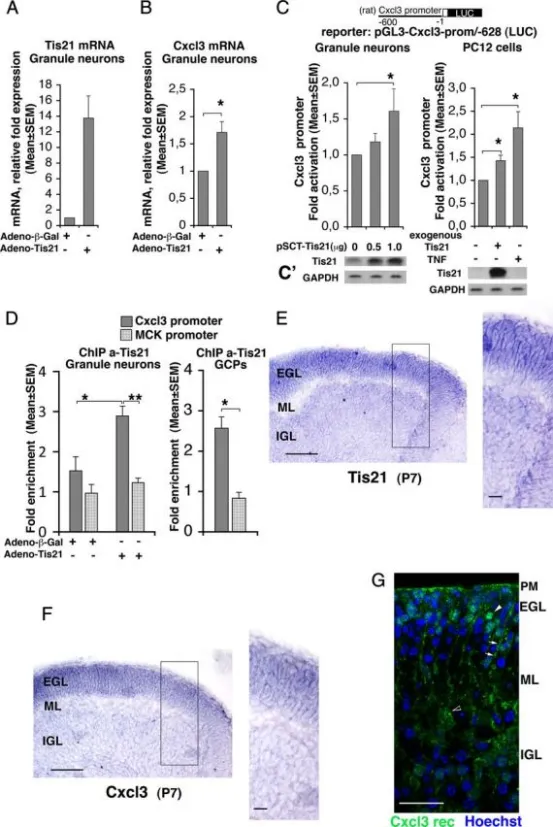
Tis21与Cxcl3启动子结合并诱导Cxcl3转录。
A、B:实时RT-PCR检测小脑颗粒神经元原代培养中Tis21与Cxcl3 mRNA的表达。细胞取自P7大鼠,种植当日(DIV 0)即感染重组腺病毒adeno-Tis21或对照adeno-β-Gal,24 h后收集。所用引物(大鼠序列)可同时检测内源和外源Tis21 mRNA。数据为5次独立实验的均值±SEM,内参为TATA结合蛋白mRNA。*p<0.05 vs对照,Student’s t检验(在以内参归一化、尚未计算为相对表达倍数的数据上进行)。
C:在P7野生型大鼠小脑颗粒神经元中转染Tis21表达质粒(pSCT-Tis21),以及在可诱导表达Tis21(大鼠序列)的PC12细胞中,Cxcl3启动子活性均显著高于仅表达内源Tis21的对照。Cxcl3启动子构建体包含转录起始位点5′端上游628 nt,克隆于荧光素酶报告基因上游(构建体pGL3-Cxcl3-prom/-628)。TNF-α作为已知Cxcl3启动子诱导剂,在PC12细胞中用作阳性对照。荧光素酶活性以相对对照的倍数表示,数据为5次实验的均值±SEM。*p<0.05 vs无外源/异位Tis21的对应对照,Student’s t检验。
C′:平行培养物以Western blot检测Tis21蛋白表达。
D:染色质免疫共沉淀(ChIP)检测Tis21与Cxcl3启动子及肌酸激酶启动子(MCK,阴性对照)的结合。实验对象:左侧柱状图所示为按A、B条件感染adeno-Tis21或adeno-β-Gal的小脑颗粒神经元;右侧柱状图所示为P7野生型大鼠小脑前体细胞(GCPs)。使用抗Tis21抗体A3H进行免疫沉淀,所得Cxcl3或MCK启动子片段量以实时PCR定量,并以富集倍数表示(Tis21免疫沉淀所得启动子百分比与正常血清对照之比)。Tis21蛋白与Cxcl3启动子的结合(深灰色柱)显著高于基线水平(即与阴性对照MCK启动子的结合水平),且与Tis21 mRNA的过表达正相关(参见A)。数据为小脑颗粒神经元5次、GCPs 3次独立实验的均值±SEM。*p<0.05 vs对照,Student’s t检验。
E、F:P7野生型小鼠小脑矢状切片原位杂交显示Tis21与Cxcl3 mRNA表达,二者在整个外颗粒层(EGL)宽度内共定位且表达极高。黑框示高倍区域。标尺:100μm;放大图20μm。
G:Cxcl3受体Cxcr2的免疫染色(特异性抗体)在P7野生型小鼠小脑矢状切片的代表性共聚焦图像。细胞核以Hoechst 33258标记。Cxcr2受体簇在GCPs(EGL内白色箭头)明显可见,软脑膜(PM)及浦肯野细胞(黑白箭头)亦有较低表达;白色箭头指示疑似Bergmann胶质细胞。标尺25μm。
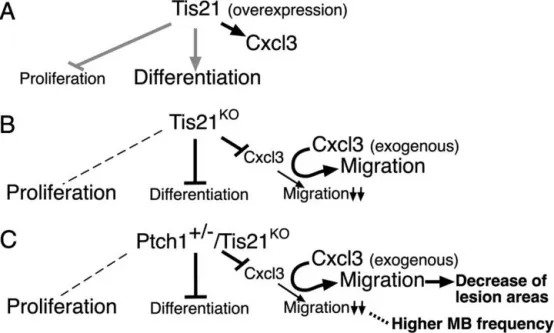
工作模型:在Tis21功能获得(A)或Tis21功能缺失(B:Ptch1+/+背景;C:Ptch1+/−背景)条件下,图示均以GCPs为对象,描述其增殖、分化以及离开外颗粒层(EGL)的迁移。A中灰色线表示来自既往文献的数据(Canzoniere等,2004;Farioli-Vecchioli等,2007)。B、C:在Tis21KO及Ptch1+/−/Tis21KO小鼠中,GCPs的Cxcl3表达减少,且GCPs迁出EGL的能力下降。C:在Ptch1+/−/Tis21KO小鼠中,上述下降与髓母细胞瘤发生率增高相关(虚线)。两种基因型中,外源性Cxcl3均可挽救迁移缺陷;在Ptch1+/−/Tis21KO小鼠中,Cxcl3还能减小小脑病变面积(C)。左侧虚线表示对增殖无显著影响(与对照相比)。
关于作者
小儿神经外科,专注于解决儿童诸多神经外科疾病,诸如脑肿瘤、脑积水、小儿癫痫、先天畸形等,一直被称为难度最大的外科专业之一。Concezio Di Rocco教授曾受到来自法国、埃及、吉尔吉斯斯坦等国家的邀请进行讲座和现场演示,且联合全球知名小儿神外专家创办了儿童神经外科研究所课程。
该项课程在过去30年里为世界培养了无数儿童神经外科医生,包括目前国内若干知名三甲医院的儿童神经外科主任专家,都曾是他的学生。Concezio Di Rocco教授将其领先的治疗技术分享给世界各地儿科神经外科医生进行借鉴参考和学习,为世界儿科神经外科的发展做出了巨大贡献。Di Rocco教授曾到访苏州大学附属儿童医院,为6名神经外科疑难杂症孤儿进行义诊。


Di Rocco教授一生撰写或参与撰写的儿童神经外科部分著作

国际儿童神经外科学会主席(ISPN)(1991-1994年)
世界神经外科联合会教育委员共同主席(2013-2017年)
世界神经外科联合会儿童神经外科委员会主席(2001-2009年)
世界小儿神经系统知名杂志《Child´s Nervous System》现任主编
国际儿童颅底学会主席(创始成员)
世界神经外科联合会基金小儿脑积水项目负责人
德国汉诺威国际神经科学研究所儿科神经外科主任(2013年至今)
欧洲神经外科学会(EANS)前副主席
世界神经外科学会联合会(WFNS)儿科委员会曾任主席
罗马天主教大学医学院儿科神经外科学系主任
国际儿童神外专家Concezio Di Rocco教授
自2014年5月起,Di Rocco教授就担任了德国汉诺威国际神经科学研究所(INI)的儿科神经外科主任。从事儿童神外事业50余年,Di Rocco教授尤为擅长小儿神经纤维瘤、癫痫、脑积水、蛛网膜囊肿、颅缝早闭、脑和脊髓肿瘤、脑和脊柱畸形(半椎体畸形,皮质发育不良,脊髓脊膜膨出,脊髓内脂肪瘤,Arnold-Chiari畸形等)难治病症方面的治疗,曾进行过12000多次神经外科手术。
- 文章标题:【INC国际大咖研究成果】Tis21敲除通过抑制Cxcl3依赖性小脑神经元迁移,提高Patched1杂合子小鼠髓母细胞瘤
- 更新时间:2025-09-08 14:10:55